|
Le gène égoïste
Depuis lintroduction des nouveaux paradigmes de la sociobiologie, on se mit à expliquer les comportements animaux (et même humains!) non plus par l'avantage de l'organisation pour le groupe (sélection de groupe), mais par la théorie des « gènes égoïstes » « qui ont inventé des artifices, les individus, pour les reproduire ». Allant plus loin, comme on le verra dans le présent chapitre, on s'est rendu compte que même les séquences sont «égoïstes »: c'est l'ADN lui même qui est «égoïste».
A présent nous avons du génome l'image d'un microcosme agité de conflits à mort, peuplé de parasites omniprésents, souvent silencieux, dont on ignore s'il sont parfois utiles, et de quelques rares gènes qui seraient «normaux », utiles au développement ou au fonctionnement de l'individu. Encore ces gènes eux mêmes sont ils mités par de longues séquences non codantes, les introns, et donc faits de morceaux qu'il faut rabouter pour les lire.
Les
rétrotransposons
Les gènes, sont des fragments d'ADN qui sont les formules de protéines. Seulement voila 97% de notre génome est muet. C'est-à-dire qui ne code aucune protéine, on l'appelle "ADN junk" ou ADN égoïste. La formule la plus commune de l'ensemble du génome est le gène qui code une enzyme : la transcriptase inverse. Il s'agit d'un gène qui ne sert a rien, mais il est vital à un certain type de parasite : les rétrovirus. Ce sont des virus à ARN comme le virus du SIDA, cette enzyme traduit l'ARN du virus en ADN et l'insère dans le génome de la victime ainsi infecté, or c'est l'un des gènes les plus courants du génome. Il existe à des centaines d'exemplaires. Cette enzyme copie aussi un gène en un brin d'ARN puis elle recopie cette dernière en la version ADN initiale qu'elle recolle dans le génome ; c'est ainsi que certains gènes de virus peuvent se recopier à foison. S'il y plusieurs copies du gène de la transcriptase inverse c'est que des rétrovirus les y ont introduits. Le génome comprend actuellement plusieurs génomes viraux presque complets dont la plupart sont inactifs, ces rétrovirus endogènes forment 1.3 % de notre génome. Certains génomes viraux n'infectent plus les personnes en sautant d'un individu à un autre par contact ou par salive mais en passant par le génome au fil des générations. Ce sont de véritables parasites génétiques. De tel rétrotransposons sont d'ailleurs plus courants que les rétrovirus. Le plus fréquent s'appelle LINE-1, il s'agit d'un paragraphe d'ADN de plus de 1000 bases qui détient en son centre la formule de la transcriptase inverse, ainsi un LINE-1 peut se transcrire lui même, fabrique sa propre transcriptase inverse, se servir de celle ci pour obtenir une copie ADN de lui même puis l'insérer dans le site de son choix. LINE-1 est très fréquent (plus de 100000 répliques) il représente 14% de notre génome, 5 fois plus que les gènes véritables. Plus fort, le parasite LINE-1 est lui même parasité, c'est ainsi qu'on trouve un paragraphe encore plus courant que LINE-1 appelé ALU (100000 de copies). Chaque ALU se réplique en utilisant la transcriptase inverse des autres et bizarrement lintron ALU ressemble au gène qui code le ribosome c'est-à-dire l'enzyme qui fabrique les protéines. Ce gène contrairement aux autres, comprend un promoteur interne, c'est-à-dire que son message "lisez moi" s'écrit dans une séquence au milieu du gène, ce qui lui permet de proliférer dans la mesure ou il porte le signal de sa propre transcription.
La guerre des sexes
Pourquoi les deux chromosomes sexuels sont-ils souvent de tailles si différentes? Dans l'espèce humaine, par exemple, l'Y (présent chez les hommes) est très petit, tandis que l'X (un exemplaire chez les hommes, deux chez les femmes) sont nettement plus grand.
Nos ancêtres, à un moment donné perdirent lhabitude reptilienne de déterminer le sexe par la température de luf pour lui substituer la détermination génétique. La raison vraisemblable à ce changement était que chacun des sexes pouvait commencer à se spécialiser lors de la conception. En ce qui nous concerne (nous humains), la détermination sexuelle génétique donnait un mâle, son absence une femelle. Le gène attira bientôt à lui dautres gènes bénéfiques aux mâles : disons les gènes dune plus grande force musculaire ou de lagressivité. Mais dans la mesure ou ces gènes là nétaient guère appréciés des femelles, des gènes secondaires se sont révélés un avantage chez un sexe et un handicap chez lautre. On les appelle les gènes sexuels antagonistes. Le dilemme fut résolu quant un autre gène mutant supprima le processus normal déchange de matériel génétique entre les 2 chromosomes appariés (crossing over). Les gènes antagonistes pouvaient désormais diverger et aller chacun leur chemin. Cest ainsi quune paire de chromosomes de taille moyenne, qui jadis accueillaient toutes sortes de gènes, fut cooptée par le processus de détermination sexuelle et devint la paire des chromosomes sexuels, chacun deux capturant des assortiments de gènes différents. Les gènes avantageux pour les mâles mais pas pour les femelles saccumulèrent sur les chromosomes Y ; les gènes profitables aux femelles mais délétères pour les mâles, sur le chromosome X. On vient par exemple de découvrir un gène baptisé dax, sur le chromosome X. Quelques rares individus naissent avec un X et un Y mais avec 2 exemplaires de dax, sur le X. Le résultat, cest que ces personnes ont beau être génétiquement des hommes, elles deviennent des femmes en tous points normales. La raison cest révélée que dax est sry (le gène situé sur le chromosome Y) qui transforme le male génétique en homme) sont antagonistes. Un sry a le dessus sur un dax, mais deux dax sont plus fort que un sry. Pour filer la métaphore on voit déjà chacun de nos 2 chromosomes cesser davoir à cur les intérêts de lespèce.
Supposons, par exemple quun gène apparaisse sur le chromosome X spécifiant la formule dun poison mortel pour les spermatozoïdes porteurs du chromosomes Y. Lhomme qui détiendrait un tel gène naurait que des filles. Toutes ses filles seraient porteuses du gène, alors que sil avait eut également des fils, aucun naurait eu le gène. Ce qui fait que ce gène devient 2 fois plus fréquent dans la nouvelle génération que sil sétait transmis normalement. Il se répandrait très rapidement. Un tel gène existe chez le papillon Acrea encedon, le rapport entre les sexes est passé à 97% de femelles. Il ne sagit que de lun des nombreux cas de cette forme de conflits évolutionnistes, baptisée « offensive du chromosome sexuel » (sex chromosom drive). Les femmes ont 2 X et les hommes 1 X et 1 Y, autrement dit les 3/4 des chromosomes sexuels sont des X et 1/4 des Y, c'est-à-dire que X a 3 fois plus de chance de faire feu sur Y que linverse. Le résultat, cest que le chromosome Y sest délesté du maximum de gènes possible tout en réduisant les autres au silence, afin de « se mettre à labri ». Cest ainsi que le chromosome Y humain a désactivé la plupart de ses gènes. Il comprend une petite zone qui semble sêtre détachée assez récemment du chromosome X, la région dite pseudo-autosomale (voir fig. 1), puis dun gène dune immense importance, le gène sry. Cest ce gène qui déclenche toute la cascade des événements conduisant à la masculinisation.
Le gène sry est spécial. Sa séquence est remarquablement stable dun homme à lautre, il est dépourvu dallèles. En revanche notre sry est très différent de celui du chimpanzé. La mesure de la distance génétique entre espèces révèle un taux de variations pour ce gène 10 fois plus important que pour les autres gènes. La raison est dans le processus de fuite qui accéléra son évolution rapide appelé balayage sélectif. Il arrive quun gène offensif apparaisse sur le chromosome X, qui attaque sry, il y a immédiatement avantage sélectif en faveur des rares sry mutants suffisamment différents des autres pour ne pas être reconnus. Le mutant commence à se répandre dans la population mâle.
Le chromosome Y nétant jamais présent chez la femme il devient libre dacquérir des gènes très défavorable aux femelles, du moment quils offrent un avantage au mâles. Chez la drosophile, le sperme est constitué de spermatozoïdes qui évoluent dans un liquide appelé liquide séminal, celui-ci contient des protéines (le produit des gènes). Pour William Rice, de luniversité de Santa Cruz, quant les drosophiles copulent ces protéines pénètrent dans la circulation sanguine de la femelle et migre, entre autres, vers son cerveau. Là, elles ont pour effet datténuer lappétit sexuel de la femelle et de précipiter lovulation. Ainsi le mâle manipule la femelle de sorte quelle renonce à saccoupler avec dautres mâles. Dans l autre camp, la pression sélective incite la femelle à résister de mieux en mieux à la manuvre. Rice a conçu une expérience pour tester se thèse. Il a empêché les drosophiles femelles de développer une résistance sélective pendant 29 générations. Dans le même temps il mettait les mâles en situation de produire des protéines du liquide séminal de plus en plus efficaces en les confrontant à des femelles de plus en plus résistantes. Au bout de 29 générations, il remit face à face les 2 lignées initiales. Le résultat fut que le sperme du mâle savait si bien manipuler le comportement des femelles quil en été devenu mortel pour les femelles.
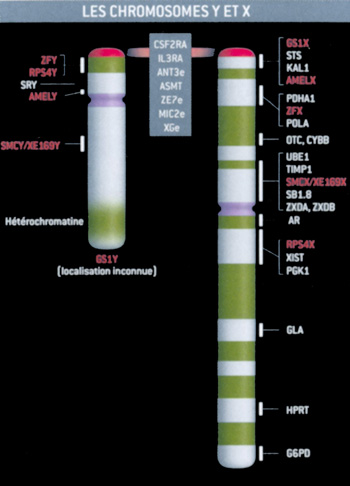 Figure
1. Quels rapports entre le X (à droite) et l'Y(à gauche) ? Les
chromosomes sexuels chez les humains n'ont en commun que quelques
gènes. La plupart constituent la région dite « pseudo-autosomale»
des chromosomes (tout en haut). Elle permet leur appariement pendant
la formation des spermatozoïdes. Le chromosome Y se termine par un
bloc d'hétérochromatine, de taille variable selon les individus.
(Source: M.A. Jobling et C.
Tyler-Smith, Figure
1. Quels rapports entre le X (à droite) et l'Y(à gauche) ? Les
chromosomes sexuels chez les humains n'ont en commun que quelques
gènes. La plupart constituent la région dite « pseudo-autosomale»
des chromosomes (tout en haut). Elle permet leur appariement pendant
la formation des spermatozoïdes. Le chromosome Y se termine par un
bloc d'hétérochromatine, de taille variable selon les individus.
(Source: M.A. Jobling et C.
Tyler-Smith,
« The Y chromosome and human evolution», TIG, 11,
449, 1995; C.M. Oisteche,
« Escape from X
inactivation in human and mouse », TIG, 11, 1, 1995.)
Les gènes
parasites
Les chromosomes sexuels n'ont pas le monopole de la bizarrerie. Ils sont même moins bizarres que d'autres. On les retrouve du moins dans chaque individu d'une espèce et leur transmission est assez régulière. Il n'en va pas de même pour les chromosomes dits « surnuméraires », les chromosomes B. Ceux-là sont présents en nombre très variable suivant les individus, et même d'une cellule à l'autre chez le même individu. Il n'y en a pas dans l'espèce humaine, mais ils sont courants chez les plantes ou les insectes. Certains criquets en ont des dizaines. De petite taille, ils portent parfois quelques gènes, certes pas indispensables, sans quoi leur absence provoquerait la mort de leur hôte. Leur seul effet global connu est de réduire la vigueur des individus. Comme on le soupçonna très tôt, ce sont des chromosomes parasites. Leur mode de transmission en témoigne.
Un individu diploïde produit des gamètes haploïdes, qui possèdent donc moitié moins de chromosomes. Normalement, les deux jeux de chromosomes que possède le parent sont représentés équitablement dans les gamètes. L'homme, qui porte un X et un Y, produira 50% de gamètes avec un X et 50% avec un Y. Cette séparation de l'information s'appelle « ségrégation ». La ségrégation la plus couramment observée est équitable, on la qualifie de «mendélienne». Mais les chromosomes B ne respectent pas la règle. Pourquoi ? Ils sont parfois présents en nombre impair, ce qui complique singulièrement le jeu de l'hérédité mais n'est pas rédhibitoire. Ce que l'on constate, c'est que les chromosomes B « trichent » lors de la méiose. Ainsi les individus portant un seul chromosome B devraient produire moitié de gamètes avec un chromosome B et moitié sans. Or on observe couramment un excès de gamètes avec chromosome B. Cette « distorsion » de ségrégation manifeste la capacité de certains chromosomes, et même de certains fragments de chromosome, à augmenter leur fréquence sans que cela soit d'aucun bénéfice pour l'organisme. Au contraire, l'individu porteur est plutôt moins vigoureux, et sa descendance (valeur sélective) est moins nombreuse. Or, par définition, un chromosome, ou une séquence qui possède l'aptitude à causer plus de distorsion de ségrégation, pourra se répandre dans une population, même si ce phénomène est ralenti par la perte de vigueur des individus porteurs.
Force est donc de revenir sur une idée fortement ancrée dans l'esprit de beaucoup de biologistes. La « fonction », plus proprement, la cause de l'existence d'une séquence présente dans un génome peut être uniquement sa seule réplication: la sélection naturelle peut favoriser ce qui améliore les individus, elle peut aussi favoriser ce qui se réplique à leurs dépens, en causant des effets délétères, si cela permet à la séquence de se reproduire mieux. Le monde des gènes a aussi ses parasites.
Parmi les chromosomes B, il en existe un que son découvreur, John Werren, n'hésite pas à qualifier de « parasite absolu ». Il se rencontre chez Nasonia vitripennis, un hyménoptère. Le déterminisme du sexe chez Nasonia, comme chez d'autres hyménoptères, est lié au niveau de ploïdie: les femelles sont diploïdes, tandis que les mâles proviennent de la ponte d'oeufs non fécondés, donc haploïdes.
Les femelles sont fécondées en général une fois dans leur vie, et serrent les spermatozoïdes dans leur réserve prévue à cet effet, la spermathèque. Pour produire un descendant mâle, elles laissent le conduit de la spermathèque fermé, c'est donc une reproduction sans fécondation (parthénogénétique). Pour produire une femelle, elles ouvrent le sphincter de leur spermathèque, l'oeuf est dans ce cas fécondé et donnera naissance à une femelle. Les mâles n'ont donc normalement pas de père, et tous les enfants d'un mâle sont normalement des femelles. On a pu reconstituer avec une bonne vraisemblance l'extraordinaire histoire évolutive qui a mené à cette bizarrerie, qui illustre la « guerre des chromosomes ».
A un moment donné de cette histoire mouvementée, les chromosomes hérités paternellement chez les mâles ont été inactivés, puis éliminés, par l'effet d'un gène particulier. C'est probablement ce gène qui resurgit dans certaines populations de Nasonia vitripennis, sur un chromosome B particulier, nommé Psr (pour Paternal Sex-Ratio). Il va considérablement perturber le déterminisme du sexe, à son grand avantage. Présent uniquement chez certains mâles, il est en fait transmis de père en fils, avec une sex-ratio de 100 % de mâles dans la descendance. Ce résultat très particulier est obtenu par hétérochromatinisation et destruction des chromosomes hérités paternellement. Il se retrouve ainsi associé à un nouveau jeu de chromosomes venu de la femelle, dans un individu mâle - puisque haploïde (fig. 2). Tout jeu de chromosomes qui se trouve associé au chromosome Psr sera détruit à la génération suivante. Psr se retrouve dans l'ensemble de la descendance d'un mâle porteur du chromosome. Il détruit toute information avec laquelle il s'associe, ce qui permet sa transmission à un maximum d'individus. Voilà une séquence pour laquelle il serait difficile de trouver une autre fonction que sa propre réplication. Le parasite absolu.
Les processus de reproduction « égoïste », on l'a dit, ne sont pas réservés aux chromosomes. Ils peuvent concerner de simples séquences, et cela nous incite à penser qu'il s'agit d'une propriété générale des séquences du chromosome eucaryote. Quand on y réfléchit, la stratégie consistant à transmettre son information aux dépens des individus qui la portent est d'ailleurs tout ce qu'il y a de plus banal: qu'est-ce qu'un mâle, sinon un individu qui insère, à l'aide d'un spermatozoïde, son information dans un gamète femelle qui fait tous les frais du développement du zygote ainsi produit? Même hors du monde eucaryote, qu'est ce qu'un plasmide bactérien, sinon une unité d'information qui se transmet de bactérie en bactérie par conjugaison bactérienne (on qualifie d'ailleurs de mâles les bactéries qui portent un plasmide...), portant principalement les gènes qui permettent ce transfert (et éventuellement quelques autres) ? Comme l'a dit Wilson, l'organisme n'est que le vecteur de transmission des gènes qu'il abrite.
Si l'on considère l'ensemble du génome eucaryote, on rencontre une foule de séquences dépourvues de fonction apparente qui peuvent donc être interprétées comme des parasites - dont l'unique fonction serait de se répliquer. Comme on le verra, il n'y a rien de plus ardu que de démontrer qu'une séquence n'a pas d'utilité. Il est presque toujours possible d'objecter que la fonction existe mais n'est pas encore connue. Et l'argument selon lequel favoriser sa propre réplication est une « fonction » suffisante pour justifier l'existence d'une séquence reste assez mal admis.
Parmi les bons candidats à l'inutilité, il y a les séquences de l'hétérochromatine constitutive. Ce sont essentiellement des séquences répétées à la queue leu leu. On peut en trouver aussi dans tous les chromosomes, spécialement près du centromère. D'une espèce à l'autre, même dans des groupes très proches, la quantité d'hétérochromatine peut varier dans des proportions considérables. Chez les grenouilles Pleurodema thaul et P. bracbyops, on observe des variations considérables de la quantité d'hétérochromatine, associées à des variations d'un facteur 3 de la quantité d'ADN. Chez les plantes, on connaît quatre espèces très proches de Crépis (Asteraceae), qu'un bon botaniste sait reconnaître, et dont le caryotype ne se distingue que par des variations de quantités d'ADN associées à des variations de la quantité d'hétérochromatine. On pourra toujours arguer qu'il existe peut-être, dans le mode de vie de ces espèces, une différence qui favoriserait les individus possédant plus d'hétérochromatine. Mais cela reste à démontrer.
Dans d'autres espèces, il est tout à fait douteux que la plupart des séquences répétées aient une fonction quelconque dans les cellules somatiques. Et pour cause: elles en sont exclues au cours des premiers stades de développement de l'embryon. Cette exclusion de larges pans du génome fut décrite dès 1887 par Boveri chez un ver parasite de l'intestin, et retrouvée chez plusieurs autres espèces d'Ascaris. Elle ne prouve pas bien sûr l'absence totale de fonction des séquences en question elles pourraient très bien avoir un rôle limité à la lignée germinale. Mais, là encore, il faudrait l'établir.
Chez l'homme, les séquences hautement répétées forment pour une large part de l'ADN dit «satellite ». Lorsqu'on classe les fragments d'ADN d'un génome par ordre de densité, apparaissent d'une part un pic central, correspondant à la teneur moyenne de l'ADN en bases GC, plus « lourdes », et d'autre part des pics décalés, «satellites», signalant des séquences nombreuses avant le même contenu en bases. Ces séquences, dont les répétitions (environ 100000 répétitions pour chacun des types de satellite) peuvent s'étendre au total sur plus de 1 million de bases, constituent 5 % de notre génome... mais 45 % de celui du cheval...
Cet ADN satellite s'accumule à proximité du centromère de quelques chromosomes. Cela indique-t-il une fonction mécanique particulière au niveau du centromère? L'hypothèse a été suggérée; elle ne repose sur aucune donnée précise. Les recombinaisons sont rares dans cette zone; or ces événements diminuent le nombre de répétitions d'une séquence. Serait-ce une raison de l'accumulation des séquences satellites? De fait, celles-ci sont nombreuses aussi dans le chromosome Y, qui ne recombine pas.
Pour voir si l'hétérochromatine sert à quelque chose, des chercheurs ont manipulé des régions hétérochromatiques chez la drosophile (délétion, translocation avec d'autres séquences). Mais les effets obtenus peuvent être diversement interprétés. En effet, quand des gènes se retrouvent au voisinage d'une région hétérochromatique dont ils sont normalement éloignés, leur transcription est ordinairement réprimée. Ce n'est pas l'hétérochromatine qui réprime directement, c'est plutôt qu'elle est repérée par un système de répression, dont l'action s'étend dans les zones voisines. Il existe des séquences plus petites, elles aussi répétées en tandem, mais dont les unités de répétition sont nettement plus courtes (une centaine, voire seulement quelques dizaines de paires de bases) : les minisatellites. Les plus célèbres sont sans doute les minis satellites des télomères (extrémités des chromosomes). Ce sont des suites de séquences de six bases (TTAGGG) chez l'homme et quelques autres espèces. Elles ont clairement une fonction: en créant une conformation particulière de l'ADN au bout du chromosome au moment de sa réplication, ou même simplement par leur répétition, elles permettent aux télomères d'être conservés tels quels au cours des mitoses successives. D'autres minis satellites, les minis satellites hypervariables, se présentent sous forme de blocs de moins de 20 milliers de bases, comprenant des répétitions de motifs du genre GGGCAGGAXG (X représentant n'importe quelle base). On connaît aussi des équivalents beaucoup plus courts, les microsatellites, répétitions de quelques dizaines de AC, de CT, ou même juste de A. La fonction de ces séquences très variables (qui fournissent donc des marqueurs d'une grande utilité en génétique) reste inconnue. Dans certains cas, elles n'en ont clairement pas comme on peut l'observer chez l'homme.
Les graves maladies génétiques dites « à expansion de triplets » sont dues à une expansion de microsatellites particuliers, le plus souvent des polymères de CAG ou CGG, ou d'autres triplets de bases. On connaît maintenant onze maladies dégénératives du système nerveux et quelques anomalies développe mentales (comme la poly syndactylie) causées par l'expansion de ces microsatellites. Certaines sont à expression dominante (amyotrophie spinobulbaire, maladie de Steinert, chorée de Huntington), les autres à expression récessive (ataxie de Friedreich, syndrome de retard mental avec X fragile). La famille des malades connaît un syndrome d'anticipation de la maladie: les personnes ayant les allèles les plus longs ont dans leur descendance des allèles de plus en plus longs. Comme la gravité et la précocité de la maladie sont proportionnelles à l'excès de taille du microsatellite, les symptômes empirent de génération en génération.
D'après l'exemple de ces maladies, il semble qu'un type de séquence particulière, ici une suite de CAG ou de CGG, ait la particularité de causer spontanément l'adjonction de nouvelles répétitions. Bien que l'on manque de données pour des séquences plus importantes, le processus est peut-être généralisable: si jamais une séquence possède une capacité plus grande qu'une autre à se dupliquer à côté d'elle-même, elle pourrait engendrer des suites de séquences répétées, a priori sans utilité aucune. Si l'on suit le raisonnement, l'encombrement dû à l'accumulation de ces séquences peut devenir un problème... à moins que des systèmes de régulation n'acquièrent la capacité de les reconnaître et de les compacter sous forme d'hétérochromatine. Un tel système existe bien chez la drosophile, où une séquence répétée plus de trois fois en tandem est compactée et inactivée. Resterait alors aux régions adjacentes à s'adapter à la présence de ces masses d'ADN réprimées dans leur voisinage, et on obtiendrait la situation observée à l'heure actuelle.
Un tel scénario semble confirmé en partie par la structure des séquences répétées de l'ADN satellite: on y retrouve souvent des répétitions emboîtées, les grandes unités de répétition contenant souvent de plus petites unités imparfaitement répétées, comme si une mutation dans une suite de répétitions d'un microsatellite pouvait créer le motif d'un mini satellite, qui pourrait se répéter à son tour (et dans lequel une mutation pourrait mener à la constitution d'une plus grande unité de répétition, etc.). Au niveau du phénotype, la seule limitation viendrait de contraintes sur le temps de développement: dans des espèces où il doit être assez court pour des raisons écologiques, la sélection jouerait contre l'augmentation de la quantité d'ADN.
D'autres séquences répétées forment aussi une part non négligeable du génome. Ce sont les transposons, séquences répétées dispersées et parfois mobiles, qui représentent chez l'homme 20 % de l'ADN total (voir les rétrotransposons ). Certaines de ces séquences sont interprétées comme apparentées aux rétrovirus. Il en est qui se comportent comme de véritables virus intra génomiques, tandis que d'autres sont devenues silencieuses: elles ont perdu la capacité de se répliquer à d'autres endroits du génome, ou même ont été « récupérées » dans des gènes fonctionnels. On connaît ainsi une protéine virale qui a été récupérée et qui joue un rôle clé lors de la fusion des membranes du spermatozoïde et de l'ovule: bel exemple de reconversion, de passage du plus extrême « égoïsme » réplicatif, celui du virus, à une contribution active à la reproduction des individus.
Et le reste? Si 65 % du génome humain est bien constitué de séquences uniques, on sait aussi qu'il ne comprend pas plus de 20 % de gènes (en comptant les introns, séquences non codantes à l'intérieur des gènes). Au total, plus de 45 %, soit près de la moitié du génome humain, sont donc constitués de séquences uniques non codantes. Cet ADN poubelle (les Anglo-Saxons disent junk) apparaît comme un ramassis de ratages de réplication, de transcription reverse (recopiage en ADN d'une information sous forme d'ARN), de vieilles séquences répétées percluses de mutations, etc.
Le génome eucaryote est finalement un bien curieux édifice. C'est moins un chef-d'oeuvre d'optimisation qu'un assemblage de séquences aux intérêts très divers, parfois divergents. Plutôt que l'image monolithique d'une gigantesque machine contribuant à ce but magnifique que seraient la création et la reproduction d'un organisme, nous voyons une société disparate d'individualismes forcenés, sanctionnés chacun par la dure loi de la sélection naturelle: seuls sont retenus ceux qui auront su se répliquer. Ce qui peut être obtenu soit en contribuant au succès de l'individu - la raison d'être d'une séquence sur cinq -, soit en parasitant l'ensemble de l'édifice, plus ou moins violemment, plus ou moins silencieusement ce qui semble être le lot du reste des séquences.
Les génomes procaryotes sont plus optimisés. Ils possèdent bien quelques séquences répétées dispersées semblables aux transposons, et des plasmides plus ou moins parasites. Certaines séquences très peu répétées (impliquées dans les « locus de contingence ») jouent un rôle majeur dans la production de variants qui échappent à nos systèmes immunitaires. Mais la majorité de l'ADN bactérien est codant. Faudrait-il voir dans la belle organisation des génomes procaryotes un trait évolué, qui les rendrait (sur ce point!) bien supérieurs aux génomes eucaryotes ? Cette idée séduit actuellement bon nombre de chercheurs.
La réussite éclatante des eucaryotes réside dans l'élaboration d'organismes complexes, grâce à une expansion considérable de leur nombre de gènes, on peut voir le génome eucaryote comme un ensemble assez mal organisé structurellement, mais qui tirerait de ce désordre une inventivité inégalable. Le gène sry , qui est le signal de départ de l'immense machinerie moléculaire menant au déterminisme du sexe mâle, semble un exemple remarquable de ce bricolage, que l'ordre bactérien aurait sans doute moins bien toléré. Ce gène en effet n'a pas d'intron, et possède des séquences typiques d'un ARN messager: en un mot, il est le résultat de la transcription reverse d'un ARN messager, un accident de fonctionnement de quelque virus ou rétrotransposon, une scorie typique du désordre eucaryote repris dans une structure fonctionnelle.
De très anciens eucaryotes, par exemple, ont un jour laissé survivre des cyanobactéries qu'elles venaient de phagocyter ; elles en ont ensuite fait de petites usines photosynthétiques. Les chloroplastes. La plupart des gènes ont déserté les chloroplastes. Pour nos mitochondries, usines cellulaires de la respiration, un phénomène identique s'est produit: on connaît même des mitochondries qui ont perdu tout génome, par transfert dans le noyau eucaryote.
On voit donc bien, à partir de ces chimères génomiques, que les différences entre les procaryotes et les eucaryotes ne portent pas principalement sur les gènes (procaryotes ou eucaryotes): beaucoup de gènes procaryotes ont pu avoir un grand succès dans des organismes eucaryotes, à l'instar des phytochromes. Pigments qui servent à l'horloge interne des végétaux et qui sont probablement d'origine bactérienne. Curieusement, l'inverse n'est pas vrai : il faut croire que l'organisation même du génome eucaryote permet cette multiplication. Cette diversification, là où la rigidité fonctionnelle et l'optimisation stricte du génome procaryote élaguent de tels édifices. Le prix en est peut-être toutes ces scories, tous ces rebuts de génomes: séquences répétées, microsatellites, hétérochromatine... Beaucoup de désordre par rapport au monde des bactéries, mais c'est peut-être le prix à payer pour l'inventivité et la complexification qui font le succès planétaire des eucaryotes!
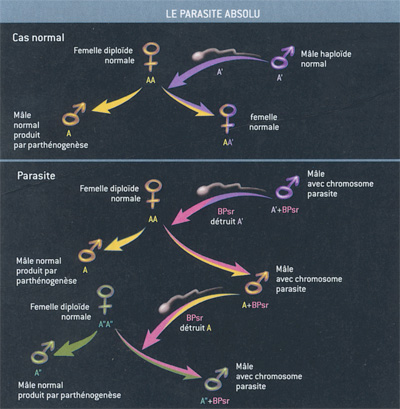 Figure 2. Le parasite absolu. Le chromosome BPsr ne
fait pas de quartier. Il tue le génome dans lequel il se trouve. Et
assure ainsi sa reproduction... Chez l'hyménoptère Nosonia
vitripennis, les chromosomes des femelles vont par paires (AA:
diploïdie). La lignée germinale des mâles est haploïde (A'). Le sexe
est déterminé par la femelle, qui peut féconder ou non les oeufs
qu'elle produit. Les mâles résultent donc d'une parthénogenèse. Psr
provoque la destruction du génome A' dans les spermatozoïdes. La
descendance d'un mâle Psr produit donc seulement des mâles Psr. Et,
à la génération suivante, le génome associé au Psr est détruit à son
tour. Il s'agit cette fois d'un génome femelle... Cela permet à Psr
de n'être présent que chez des mâles, qui le transmettent à 100 % de
leur descendance. Figure 2. Le parasite absolu. Le chromosome BPsr ne
fait pas de quartier. Il tue le génome dans lequel il se trouve. Et
assure ainsi sa reproduction... Chez l'hyménoptère Nosonia
vitripennis, les chromosomes des femelles vont par paires (AA:
diploïdie). La lignée germinale des mâles est haploïde (A'). Le sexe
est déterminé par la femelle, qui peut féconder ou non les oeufs
qu'elle produit. Les mâles résultent donc d'une parthénogenèse. Psr
provoque la destruction du génome A' dans les spermatozoïdes. La
descendance d'un mâle Psr produit donc seulement des mâles Psr. Et,
à la génération suivante, le génome associé au Psr est détruit à son
tour. Il s'agit cette fois d'un génome femelle... Cela permet à Psr
de n'être présent que chez des mâles, qui le transmettent à 100 % de
leur descendance.
La transduction
Une petite fraction des particules produites par une bactérie, à la suite de l'infection due à certains bactériophages (virus), est formée d'une capside de protéines virales entourant un fragment d'ADN bactérien. Ainsi, dans une bactérie infectée, l'assemblage des protéines virales, donc l'édification du virion, peut, accidentellement, s'effectuer autour d'un morceau d'ADN bactérien au lieu d'une molécule d'ADN du virus. Ces particules hybrides dites transductrices, peuvent, tout comme les virus normaux, infecter une bactérie sensible et y injecter l'ADN qu'elles contiennent. Les rats quittent le navire !! On voit là un phénomène bien étrange, après avoir subit l'agression de la part des gènes du virus, les gènes bactériens s'échappent et se mettent à l'abri !
Les
memes
Richard Dawkins étend l'approche Darwinienne en introduisant le concept de même (analogue mental du gène). Pour cet auteur, un même est un objet mental qui, tel une créature biologique, lutte pour sa survie. A l'instar du gène égoïste (le gène est un cas particulier de même), le même se sert de l'individu qui en est porteur afin de se disséminer. Cette dissémination n'est pas, au contraire du domaine biologique, uniquement liée à une activité de reproduction mais dépend des moyens de communication offerts à l'individu. Or la variété de ces derniers, depuis la banalisation de l'accès aux moyens de télécommunication, est de plus en plus aisée. Dawkins voit donc le champ culturel comme le siège d'une activité dans laquelle les mêmes interagissent par le biais de leurs véhicules humains. Grâce à l'expression orale, écrite ou visuelle, certaines interactions peuvent aboutir à l'extinction de mêmes (la mort subite d'un unique porteur d'un même en est un exemple extrême), d'autres à des associations entre mêmes dont l'adoption simultanée par un individu ou par une communauté en améliore la survie. Nous observons encore l'occurrence de deux concepts :
- la diversité des mêmes est à la fois la conséquence de la diversité génétique des individus et des environnements dans lesquels ils sont plongés. Des événements aussi simples que le lever et le coucher du soleil sont à la source de nombreux mythes car ils sont partagés par toute l'humanité. En revanche, les particularités de la faune, de la flore (qui résulte elle-même de la situation géographique, de la géologie) et le degré d'isolation dans lequel se trouve une population, conduit cette dernière à établir des croyances spécifiques.
- la sélection des mêmes qui peut s'effectuer de manière dramatique, par la sélection des individus eux-mêmes : un individu possédant une attitude suicidaire voit ses chances de faire des adeptes fort limitées. Plus heureusement, un ou plusieurs individus dont les traditions améliorent l'état physique ont plus de chances de procréer, d'accumuler de l'expérience et de la transmettre à leurs descendants. En cela, R. Dawkins compare le cheminement des idées dans l'esprit des hommes à celui des gènes au sein d'une espèce. En prenant une image anthropomorphique, les gènes et les mêmes sont égoïstes. Tous deux sont immortels, et utilisent les organismes dans lesquels ils résident pour leur réplication et leur dissémination.
Le système de Dawkins reprend l'hypothèse de K. Lorenz en ce qui concerne le caractère universel de l'acquisition de l'information, en l'appliquant à l'évolution de la culture humaine. Mais il va plus avant en s'inspirant des mécanismes biologiques pour expliquer la formation et la persistance de mêmes de plus en plus complexes. Dans le cadre de la construction de systèmes artificiels, cette approche est pertinente au sens où elle donne une idée de la mécanique par laquelle les modèles mentaux peuvent évoluer.
Il est important de noter, comme le font remarquer J.-P. Changeux et A. Connes, qu'au niveau du même, la pression de sélection au niveau du gène n'intervient pas nécessairement et ne suffit pas à expliquer l'immense variété du patrimoine culturel humain. Il faut alors considérer que lorsqu'un même est indépendant de la pression de sélection à laquelle l'individu ou le groupe sont soumis, sa capacité à se disséminer dépend alors de son aptitude à exploiter des niches psychologiques et non plus écologiques.
Il faut considérer qu'avec l'homme, l'apparition du langage a produit une entité comparable à une forme de vie : le même. Mais à la différence d'une forme de vie biologique, un même peut être stocké sur différents supports, transmis rapidement et dupliqué à peu de frais. Il est même possible d'appliquer la métaphore des mêmes aux hypothèses les plus extrêmes concernant l'évolution humaine. Ainsi, H. Moravec décrit des machines artificielles capables de traiter (bien qu'il n'utilise pas explicitement le terme) des mêmes produits par les humains, et capables ensuite d'en créer de nouveaux. Cet auteur utilise le terme de décollage génétique (genetic takeover) pour décrire ce processus qui ferait de la machine pensante un descendant de l'homme. Si l'on conserve l'approche de Dawkins, celle du même égoïste, la machine pensante n'est-elle pas le moyen le plus adapté à la survie de certains mêmes, notamment ceux qui concernent les machines pensantes, qu'elles soient naturelles ou artificielles?
Les
viroïdes
Les viroïdes sont de simples boucles d'ARN d'environ 300 nucléotides, qui infectent certains végétaux (pomme de terre, tomate, cocotier, etc.), ne codent aucune protéine, et dont la pathogénicité serait liée à des interactions avec les mécanismes cellulaires de la synthèse protéique (maturation des ARNm).
Ce voyage au sein du génome nous a montré l'existence d'une véritable faune génétique avec des gènes pouvant être des proies, des prédateurs et des parasites, la question qui se pose alors est : sommes nous devant un monde vivant fractal, c'est-à-dire comme les poupées russes, après avoir étudié la faune animale (pléonasme) et ces gènes égoïstes verrions-nous apparaître derrière cette faune génétique des molécules égoïstes ? Est-ce que le gène forme l'atome fondamental de la dynamique du vivant ou est-ce que on peut aller plus bas ? Le prion peut fournir un exemple de molécule égoïste, il s'agit d'un agent infectieux ne contenant aucune information génétique, le prion est une molécule de protéine aberrante, dérivée d'une molécule existant normalement dans le métabolisme. Le prion a la propriété de communiquer sa conformation aux molécules normales de même catégorie, de sorte que tout se passe comme s'il se multipliait. C'est l'agent pathogène de l'ESB et de la maladie de Creutzfeldt-Jakob.
|